
Noise Criteria
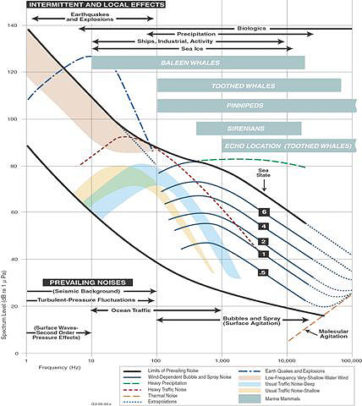 One of the most tenacious challenges in establishing mitigation guidelines for marine animal noise exposure is the lack of a clear numeric threshold that regulators can unambiguously use. There is no “55 miles per hour” in marine bioacoustics because the physical environment of the ocean and the biological adaptations to habitat are extremely complex.
One of the most tenacious challenges in establishing mitigation guidelines for marine animal noise exposure is the lack of a clear numeric threshold that regulators can unambiguously use. There is no “55 miles per hour” in marine bioacoustics because the physical environment of the ocean and the biological adaptations to habitat are extremely complex.
There is a well known need for a marine “Noise Exposure Criteria” such as is used in contemporary architectural practice. But architectural noise criteria have been established over the years through empirical assessment of how humans use and inhabit built environments, and are predicated on our priorities of speech intelligibility, annoyance, and physiological damage. And while noise criteria for animal enclosures are beginning to appear, these criteria are for domesticated animals with behaviors that we know and understand.
The range of species and adaptations in the ocean are much more complex. Some animals live in extremely noisy environments – such as intertidal zones and Arctic ice flows. Some animals need to perceive highly detailed sound characteristics, others only need to hear rudimentary thuds and bumps; some animals use sound to see and navigate, others only to threaten or court.
Given the vast repertoire of bioacoustic adaptations found in the ocean, trying to find one number or one measurement that expresses conflicting or compromising noise exposures would be impossible. But as our understanding of animal interactions with their habitat unfolds we can begin to find ways to express these interactions in well defined terms and quantitative expressions.
Thresholds
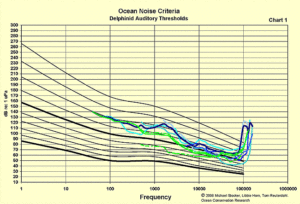 At this juncture establishing what marine animals actually can hear is a challenge. There has been some detailed work on a few captive marine mammals, and a legacy of testing on fish. But audiological tests on any animal are difficult because heretofore most testing uses what is termed “operant conditioning” which involves training or “conditioning” an animal to associate a sound with either a reward or an aggravation. Once the animal perfects the response, the sound parameters are changed in some manner to bracket the limits and thresholds of how it hears sounds which it associates with the reward or aggravation.
At this juncture establishing what marine animals actually can hear is a challenge. There has been some detailed work on a few captive marine mammals, and a legacy of testing on fish. But audiological tests on any animal are difficult because heretofore most testing uses what is termed “operant conditioning” which involves training or “conditioning” an animal to associate a sound with either a reward or an aggravation. Once the animal perfects the response, the sound parameters are changed in some manner to bracket the limits and thresholds of how it hears sounds which it associates with the reward or aggravation.
So for example a dolphin is trained to expect a reward if they correctly identify a tone, theoretically if the pitch or amplitude of the tone is modified the dolphin will seek a reward if they can hear the new tone, and won’t seek a reward if it can’t hear a tone. Through this very time consuming process the limits and extents of the dolphin’s hearing can be ascertained. And while this would be a captive, trained, and thus “domesticated” animal, there will be some correlation between this dolphin’s hearing and the hearing of its wild kin.
Aside from the training and time-consuming procedure, some other drawbacks include:
- Captive animals become habituated to testing and serve as a poor proxy for wild animals
- Some animals such as baleen whales are not easily captured, let alone trained, so we can only speculate about their hearing
- Test signals are most often calibrated laboratory signals that may not correlate to actual exposure signals
- Test signals in captive enclosures mostly occur in the near field (close proximity) and don’t accurately reflect time and spatial domain sound characteristics found in marine habitats.
- Testing individual animals that are usually schooling animals does not reflect aggregate signal amplification and behavioral adaptations that occur in group settings.
- Most testing is done using “pressure gradient” acoustical energy at a specific receiver point and does not reflect other characteristics of acoustical energy perception.
- Auditory testing typically focuses on sensitivity to how loud a signal is, and the extents of the perception of frequency or “pitch.” Other sound characteristics are rarely considered.
We have accumulated, evaluated, and plotted audiograms for marine mammals and fish. It is clear from these graphs that there is a disparity in the tested hearing of the various animals. There is more coherence among marine mammals because there are many similarities in their hearing systems. For example all mammals have a pitch discrimination organ in their inner ear called a “cochlea.” There is no analogous organ in fishes, so it is quite possible that pitch discrimination is not a useful adaptation for this class of animal.
Nonetheless audiograms for fish still represent amplitude/pitch sensitivity partially because amplitude/pitch sensitivity reflects our own auditory priorities. The lack of coherence among fish audiograms also suggests the diversity in their adaptations to their particular habitats.
Kurtosis
 There are characteristics of sounds found in nature which differ from sounds produced by humans. Marine animals have adapted to natural sounds over the eons. We know that some of the sounds produced by humans are compromising natural acoustical relationships. So focusing on the differences between natural and un-natural sounds can give us a handle on how to express potential impacts to marine acoustic habitats.
There are characteristics of sounds found in nature which differ from sounds produced by humans. Marine animals have adapted to natural sounds over the eons. We know that some of the sounds produced by humans are compromising natural acoustical relationships. So focusing on the differences between natural and un-natural sounds can give us a handle on how to express potential impacts to marine acoustic habitats.
One characteristic we are exploring is “kurtosis” which is a numerical expression of how “rough” or screechy a sound is. If you take two sounds at equal amplitude, and approximately equal frequencies – such as a bowed violin string and fingernails skittering across a blackboard, it is clear that one is sweet, and the other agonizing. “Kurtosis” expresses this quality difference in a numeric form.
Kurtosis is a statistical expression of signal variability over time. There is no constraint in the time domain, so the metric can be used across all frequencies – from high frequency communication sonars to thumping of marine pile driving. We believe that we will find a correlation between kurtosis and acoustical compromise.